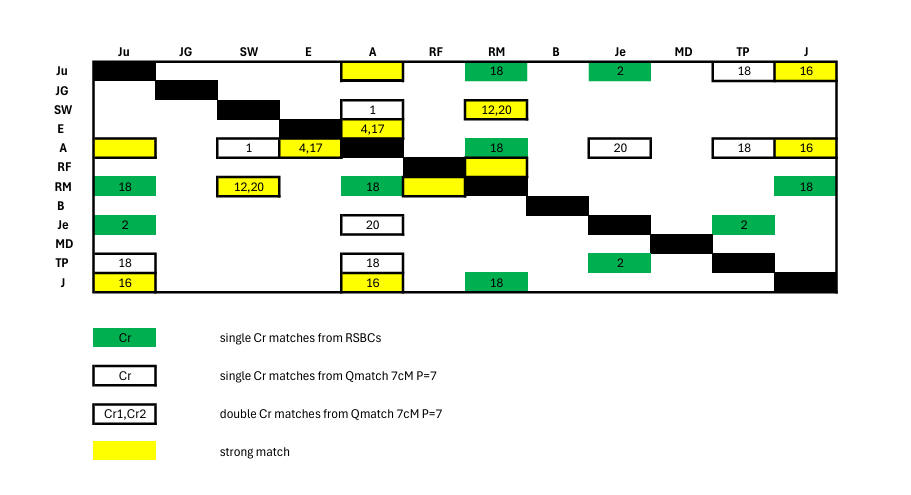
We embarked upon development of a procedure for using RSBCs to enhance proof of small matches (originally described here), because with our original set of 7 family members, there were no high-quality autosomal matches available (by which is meant 7cM with P=7 which are GEDmatch's Qmatch default settings and tough enough to deny the B/Je and TP/Je Ancestry matches which reappear here at 5cM and P=7, and the B/TP Ancestry match which reappears here at 5cM and P=3). Amazingly, with the introduction of 5 new family members - A, J, RM, RF and TP - we found a pattern of matches which was useful. In fact, as can be seen from the following diagram, it is more extensive than the matches revealed by our small match procedure using RSBCs:
The matches given by Qmatch show interbranch connection - between A and the Fischls (SW) and the NY Lousadas (Je and TP). They also show that RM matches SW. But our small match procedure adds something - Cr2 giving Ju and TP both matching Je, and Cr18 (same location as in the Qmatch) giving us RM matching the 3 Barrow Lousadas Ju, A and J. This reinforces what Qmatch tells us - namely that the English Barrow Lousadas are related to the NY Lousadas, to the Fischls and to RM. (Incidentally the Cr20 locations in 2 of the Qmatches are not the same).
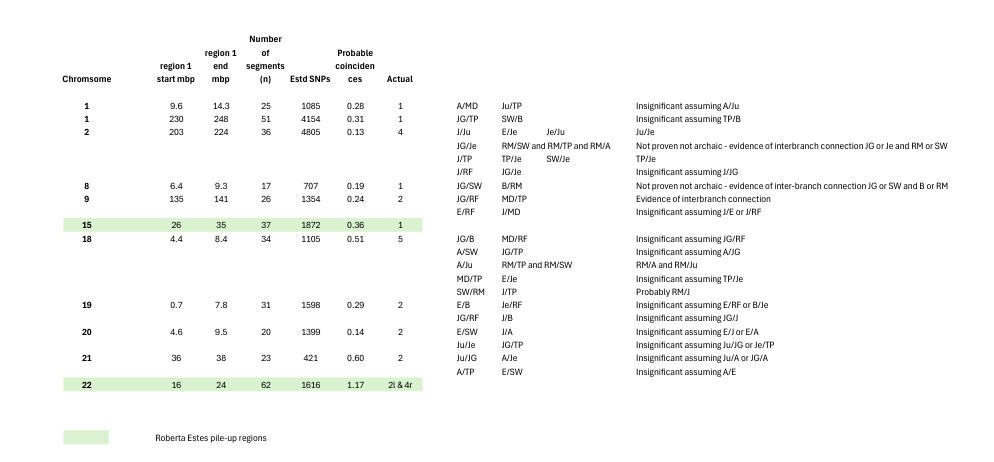
Our estimation of the random occurrence frequency for RSBCs was revised after our original description of our small match procedure. For the random occurrence needs to be estimated on a chromosome-by-chromosome basis at least - and this almost doubles the expected number of RSBCs. But crowded individual segments of chromosomes also need separate consideration - for in such regions the random occurrence is much greater than in less crowded regions. In fact, we were able to usefully classify regions according to whether the probable expected number of RSBCs expected at random was less than 0.1 or greater than 0.1.
In the less crowded regions, in all but 3 cases, genealogy rules out the RSBCs having significance, for it provides an explanation for one of the possible matches entailed in those RSBCs (see here for an explanation of what the RSBCs may entail). But in three cases there is possible significance except that we cannot be sure that the matches are not archaic. Indeed, in the Cr10 location we found 5 matches with others sharing the 135656 location as a segment boundary.
With the crowded regions, it can be seen in the following diagram that we rejected 2 well-known pile-up regions on Cr16 and Cr22. But we also rejected an obvious further case - the 752721 location on Cr1. It can be seen that many of the matches are made insignificant by genealogy. A few seem likely to be archaic. But Cr2 and Cr18 appear to be giving us genuine matches: